Range: S. Africa to Somalia and to India and Sri Lanka, absent from Red Sea and Persian Gulf; Indonesia to Philippines and to Papua New Guinea, Solomon Is. and Queensland.
Description: Moderately small to moderately large, moderately light to solid. Last whorl conical, ventricosely conical or broadly and ventricosel y conical, occasionally pyriform; outline variably convex at adapical fourth to half and straight to moderately concave below; left side often sigmoid. In Indian populations, last whorl most conical, RD most variable. Shoulder angulate to subangulate, distinctly tuberculate to irregularly edged. Spire of low to moderate height, outline slightly concave to slightly convex. Posmuclear spire whorls variably tuberculate. Teleoconch sutural ramps almost flat, with 1 increasing to 4-7 spiral grooves; spiral sculpture may be weak in last whorls. Entire last whorl with rather evenly spaced, wrinkled spiral ribs and alternating wrinkled threads; spiral sculpture may be weaker toward shoulder, and some specimens are quite smooth.
| Shell Morphometry | ||
|---|---|---|
| L | 25-64 mm | |
| RW | - | |
| RD | - | |
| PMD | - | |
| RSH | - | |
Ground colour pale greyish blue or pale pink. Last whorl spirally banded with tan to olive brown or orangish brown, usually with variably broad ground-colour bands at centre and shoulder. In Indian Ocean populations (Pl. 5, Figs. 1 -8), last whorl often maculated with bluish brown axial streaks; overlying spiral rows of brown dots sparsely to heavily developed on ribs. Basal part of last whorl and columella dark brown; in Indian shells (Pl. 5, Figs. 1,3,4), basal part mostly orange or tan, often similar to adjacent parts. Apex light brown or pink. Teleoconch sutural ramps radially maculated with orange to blackish brown, varying from sparse spots between marginal tubercles to solid dark brown colouration. In small adults, aperture bluish violet, edged with brownish violet, with a pale central band and a translucent marginal zone; aperture white in large adults.
In Indian shells, periostracum brown or yellowish brown, variably thick, slightly translucent to opaque, with about 20 variously arranged tufted spiral ridges.
Dorsum of foot brown or black anteriorly, with a red distal edge; sides of foot reddish brown, flecked with white dorsally; sole and ventro-lateral edges of foot red. Rostrum reddish brown, red distally. Tentacles light brown to red. Siphon reddish brown dorsally, edged with pale red distally (Indian animals, Kohn, 1978a & unpubl. observ.). In Indonesian and W. Pacific animals, foot and siphon brownish red dotted with black; with sparse white dots on siphon. Sole serration, of foot and siphon may also be light red, with thin transverse I brown lines on the dorsal side of siphon. Rostrum and tentacles dark red to brown (Chaberman, pers. comm., 1981; Kohn, unpubl. observ.) (Pl. 74, Fig.7).
Habitat and Habits: Intertidal and slightly subtidal. In Mozambique and South Africa, at and below low-tide level in sandy crevices and caverns (Kilbum & Rippey, 1982; Grosch, pers. comm., 1989). Some Indian populations occupy small caves and associated tidal pools on terraced intertidal limestone benches (Kohn, 1978a). In Indonesia, on intertidal to slightly subtidal benches of truncated reef limestone, on varied hard and soft substrata (Kohn & Nybakken, 1975). In N. Papua New Guinea, common on exposed shallow reefs (Chaberman, pers. comm., 1981); more frequently found on extensive mud flats of mainland habitats than on coral reef in E. Australia (Huish, 1978). The species is known to prey on eunicid and nereid polychaetes (Kohn & Nybakken, 1975; Kohn, 1978a). In Queensland, C. biliosus deposits small, stocky egg capsules underneath rocks near the reef crest (Loch, pers. comm., 1987). In Pakistan, number of eggs per capsule is 100- 1,410; egg diameter of 160- 180 µm, predicts a miminum pelagic period of 27-25 days (Zehra, 1990; Perron & Kohn, 1985; Zehra & Perveen, 1991).
Discussion: C. biliosus is most similar to C. hyaena. For the distinctions between them, see the Discussion of the latter species. Most authors regard C. panulus as a distinct, valid species. Coomans et al. (1982) provisionally separated C. biliosus, C. imperator, and ,em>C. parvulus at the species level, while da Motta (1 993) did so with C. biliosus, C. pawulus (C. imperator as synonym), and C. neoroseus (shells from Philippines and Indonesia). Here, all these nominal taxa are considered conspecific and even not conclusively separable at the subspecies level. The South Indian population is distinctive in its size and colour, but shows no clear differences in other shell characters. Walls ([1979]) described populations from South Africa and Mozambique (Pl. 5, Figs. 5, 6) as a separate subspecies (C. b. meyeri). These shells tend to be smaller than Indian shells and have a more convex outline of the last whorl, and more often weak tubercles. However, they intergrade with shells from southern India and the differences in shell morphometry are minimal. Our results suggest a clinal variation, in agreement with Kilbum (1982). C. biliosus refers to a specimen from the Gulf of Mannar, India, C. punctatus and C. piperatus seem to be synonyms. C. neoroseus is based on a specimen assumed to be from Philippines (da Motta, 1993); we know of similar shells from Indonesian populations. Specimens matching the lectotype of C. parvulus are known from Philippines and the W. Indian Ocean. C. imperator represents the Queensland population. The original figure of C. concinnus (= C. sapphirostoma) shows a small shell with smooth last whorl as occurring in Queensland and S. Africa (see Coomans et al., 1985a).
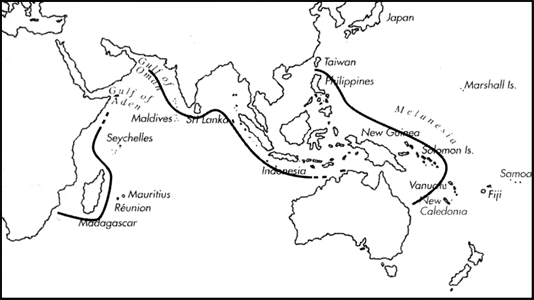
C. biliosus range map
This section contains verbatim reproductions of the accounts of 316 species of Conus from the Indo-Pacific region, from Manual of the Living Conidae, by Röckel, Korn and Kohn (1995). They are reproduced with the kind permission of the present publisher, Conchbooks.
All plates and figures referred to in the text are also in Röckel, Korn & Kohn, 1995. Manual of the Living Conidae Vol. 1: Indo-Pacific Region.
The range maps have been modified so that each species account has it own map, rather than one map that showed the ranges of several species in the original work. This was necessary because each species account is on a separate page on the website and not confined to the order of accounts in the book.