Range: Entire Indo-Pacific.
Description: Medium-sized to large, moderately solid to heavy; in similarly sized specimens, relative weight may vary by 80% between different forms and by 40% within the same form. Shells smaller in populations from the W. Indian Ocean; lightest forms also from this area. Last whorl ventricosely conical to ovate or conoid-cylindrical, sometimes also cylindrical, conical or broadly ovate; outline strongly convex to nearly straight; left side straight to strongly concave near base. Aperture variably wider at base than near shoulder. Shoulder angulate to rounded, often within the same population. Spire generally of moderate height, outline usually straight to concave. Larval shell usually of 3-3.5 whorls, of 2-2.5 whorls in some forms from the W. Indian Ocean; maximum diameter 0.7-0.9 mm. First 3-5 postnuclear whorls tuberculate. Teleoconch sutural ramps flat to slightly concave, with 0-2 increasing to 7-15 fine spiral grooves; sculpture distinct to obsolete on latest ramps. Last whorl with variably weak spiral ribs near base, occasionally followed by spiral striae to shoulder.
| Shell Morphometry | ||
|---|---|---|
| L | 40-150 mm | |
| RW | 0.10-1.30 g/mm | |
| (L 35-120 mm) | ||
| RD | 0.50-0.72 | |
| PMD | 0.67-0.86 | |
| RSH | 0.11-0.20 | |
Ground-colour white, sometimes suffused with blue, beige, violet, orange, or pink. Last whorl generally with a network of light to dark brown lines edging tiny to moderately large tents, sometimes also small quadrangular to round markings. Yellowish brown flecks and blotches arranged in 2-3 interrupted to solid spiral bands, on both sides of centre and often below shoulder; bands interspersed with broad, dark brown, straight to wavy axial lines. Occasionally, 1-2 additional similar but narrower spiral bands present basally. In the W. Indian Ocean, pattern varying from the typical arrangement to a delicate network with or without spiral bands or to a fine axially lineate pattern without any tentmarks. Larval whorls coloured with shades of red, sometimes white; colour slightly variable within the same population. In Hawaii, larval whorls changing colour from yellow to red during metamorphosis (Perron, 1980). First 2-4 postnuclear sutural ramps usually similar in colour to larval whorls, immaculate in first 1-2.5 whorls, with marginal brown dots in remaining whorls. Following sutural ramps matchng last whorl in colour pattern. Aperture usually white to bluish white, or rarely suffused with pink.
Periostracum grey to yellow, thin, translucent, smooth.
Foot and rostrum white to pale yellow or pinkish cream. Dorsum of foot mottled and netted with light to blackish brown, with a pre-marginal band of black spots ending in a lateral black blotch anteriorly; anterior part edged with orange-brown to red, with a central black blotch, sometimes also posterior end edged with red. Sole of foot mottled and transversely streaked with brown. Rostrum washed and mottled with pale brown, immaculate distally. Proboscis pale red, darker at tip. Tentacles white, lightly mottled with brown at base and tipped with red. Siphon white, tipped with red and mottled with brown at base, with a broad black ring about 1/3 of the length from the tip (Bergh, 1895; Kohn, 1959a; Walls, [1979]; Chaberman, pers. comm., 1981; Pearson, unpubl. observ.; Fainzilber et al., 1992) (Pl. 76, Fig. 65; Pl. 84, Fourth row, left).
Radular teeth comparatively long (tooth/shell length-ratio 0.11 (Kohn, 1963) to 0.29 (James, 1980)) and slender, with 2 opposed barbs adapically; long but weak to obsolete serration ending in a weak cusp about 1/7 of the length from the base; basal spur absent (Troschel, 1868; Bergh, 1895; Shaw, 1914; Peile, 1939; James, 1980; Bandel, 1984; Kohn, unpubl. observ.; Rolán, 1993).
Habitat and Habits: Intertidal to about 50 m; on coral reef from the reef crest to deeper water inside the lagoon and sometimes also on flats of mainland coasts, in sand bottoms without or with sea-weed, in coral rubble, on slabs of rock, on pinnacles of dead coral and muddy substrate, often hidden beneath rocks and coral boulders. In Mozambique, populations rather typical in colour and pattern on sand and reef substrate, while populations with shells of dark ground-colour (incl. form cholmondeleyi, see below) on bottoms of silt or muddy sand. Magnetite and manganese components of the latter substrates are supposed to be responsible for the darker colour of the shells (Cernohorsky, 1964; Kohn, 1959a, 1960, 1968b; Kohn & Nybakken, 1975; Huish, 1978; Kay, 1979; Kilburn & Rippey, 1982; Sharabati, 1984; Reichelt & Kohn, 1985; Tirard, pers. Comm., 1989; Grosch, pers. Comm., 1989; Lorenz, pers. Comm., 1993). C. textile preys on many different species of prosobranch gastropods (Conidae, incl. dangerous molluscivorous or piscivorous species as C. pennaceus or C. striatus, Terebridae, Mitridae, Nassariidae, Muricidae, Thaiididae, Coralliophilidae, Vermetidae, Siliquariidae, Cypraeidae, Turbinidae, Trochidae) and practices cannibalism after a long time of starving (Cruz, Corpuz & Olivera, 1978). Juveniles of C. textile also feed on gastropods. The radular tooth of C. textile is released immediately after injection into the foot of its prey and the proboscis is withdrawn; the stinging process may be repeated up to six times. The rostrum is then applied to the prey's aperture and the ingestion process needs about 20 minutes. However, C. textile also feeds on small fishes, worms, dead cephalopods and peeled shrimps (Kohn, 1959b, 1968b, 1983; Kohn & Nybakken, 1975; Reichelt & Kohn, 1985; Schoenberg, 1971; Cruz, Corpuz & Olivera, 1978; Perron, 1981b; Collins, 1987; Loch, pers. Comm., 1987; Tirard, pers. Comm., 1989; Grosch, pers. Comm., 1989). The venom of C. textile is toxic to polychaetes, gastropods, fishes and mice; the species is reported to have caused human fatalities. However, the results of studies on the toxicity to mammals are somewhat contradictory: Kohn, Saunders & Wiener (1960) report lethal effects in mice after intraperitoneal and intravenous injection, whereas Endean & Rudkin (1963) did not observe lethal effects in mice. According to Cruz, Corpuz & Olivera (1976), mice always recover from a intraperitoneal injection but die after an intracisternal injection. At the western side of Wheeler Reef (Townsville, Queensland), egg capsules have been collected in 18 m beneath a coral plate among brain corals. These capsules measure 31-33 x 21-26 mm each containing about 1,300 eggs, while the mean capsule size in Hawaii is 21 x 17 mm. Egg diameters of 230-270 µm (E. Indian Ocean, Philippines, Palau, Queensland, Hawaii) suggest minimum pelagic periods of 21-17 days (Huish, 1978; Perron, 1980, 1981a, b; Perron & Kohn, 1985).
Discussion: C. textile is similar to C. canonicus, C. abbas, typical C. telatus, C. victoriae, C. gloriamaris, and C. dalli. C. canonicus is separable from some C. textile variants only with difficulty. It generally has a more cylindrical last whorl with straighter sides, and its last whorl pattern is usually finer. Forms of C. textile from the W. Indian Ocean with a similarly fine reticulate pattern lack the pink aperture, while form textilinus (see below), sometimes similar in colour pattern and aperture colour, is distinguished by its ventricosely conical last whorl. C. abbas differs in having an orange rather than red or white apex and rhomboid ground-colour markings on its last whorl. Form textilinus, more similar to C. abbas than other forms of C. textile, can also be separated by its pinkish violet aperture. Only the typical form of C. telatus may be confused with C. textile. The former is generally less convex in outline and has more tuberculate postnuclear whorls (5-7 vs. 3-5), a weaker spiral sculpture on the late teleoconch sutural ramps, and finer reticulated lines. C. victoriae is dissimilar in size, shape and colour pattern from the sympatric typical form of C. textile. It differs in having smaller mean size, a broader, paucispiral larval shell, and finer reticulated lines. For comparison with C. gloriamaris and C. dalli (Vol. 2), see the Discussions of those species. C. textile is rather uniform in shape and colour pattern between and within the populations from the Central Indian Ocean to Hawaii and French Polynesia but very variable between and within the populations from the W. Indian Ocean. The vagility of the larval stage may be responsible for a pronounced intergradation between the different forms from the latter area. The variability has led to the introduction of many nominal taxa, most of which likely refer to forms, and only a few to local races or geographical subspecies. Whether some of the forms are valid sibling species of C. textile requires much further study on the soft parts and genetic analysis. Distinguishing these variants from one another, former authors mainly emphasized differences in colour pattern that, however, did not prove to be constant. Data on ecology, zoogeography and morphology of soft parts of the western populations are largely lacking, precluding any strong taxonomic hypothesis. The nominal taxa: - C. archiepiscopus (Pl. 68, Figs. 1, 4, 6): Last whorl ventricosely conical to ovate, with a fine reticulate pattern and distinct spiral colour bands. Ground colour white, sometimes suffused with blue. Known from different localities within the W. Indian Ocean. We consider it a form of C. textile very similar to form euetrios (see below). In Madagascar it intergrades with sympatric conoid-cylindrical variants of C. textile. Shells similar in shape and colour pattern sometimes occur in Melanesia. C. communis is a synonym (Coomans et al., 1985a). - C. auriger (Pl. 67, Fig. 6): Last whorl ovate, with a rather widely meshed pattern and a few yellowish brown blotches arranged in a spiral row on each side of centre. We consider C. auriger a form of C. textile hardly separable from form verriculum and intergrading with form ponderosa (see below). Richard (1990) regards C. auriger as a valid species and C. verriculum as a synonym. - C. cholmondeleyi (Pl. 67, Figs. 18-24): Last whorl conoid-cylindrical to ventricosely conical, rarely ovate; shoulder usually subangulate to rounded. Ground-colour white to brownish beige or pale violet, streaked or suffused with blue on the last whorl. Pattern consisting of 2-3 obsolete to distinct spiral colour bands and variably fine, dark brown axial lines, usually undulating but often partially reticulated. Known from Kenya to Madagascar and Natal. C. eumitus is a synonym. C. eumitus of authors (Pl. 67, Figs. 23, 24) refers to a more solid variant from Natal and Mozambique with an angulate to subangulate shoulder, a more conical and often broader last whorl, and 3 prominent spiral colour bands; the aperture may be suffused with pink. Lauer (1987) named the latter variant C. cholmondeleyi form fascifer; he emphasized differences in colouration and pattern but disregarded morphological differences. In Zanzibar, Mozambique, and Natal, C. cholmondeleyi intergrades with sympatric typically patterned forms of C. textile (Kilburn & Rippey, 1982; Grosch, pers. comm., 1989). We therefore consider C. cholmondeleyi a form of C. textile rather than a valid sibling species. - C. communis: A synonym of C. t. form archiepiscopus. - C. concatenatus (Pl. 67, Fig. 5): Likely a form of C. textile with a very reduced reticulate pattern. - C. corbula (Pl. 68, Fig. 5): A form of C. textile similar to form archiepiscopus. - C. textile f. euetrios var. cyanosus: See C. textile var. euetrios. - C. textile dahlakensis (Pl. 67, Fig. 8): Last whorl conoid-cylindrical to ventricosely conical, with a rather typical C. textile pattern. Originally described as a local race from the Dahlak Archipelago, Red Sea. Shells matching these specimens from Dahlak Archipelago also occur from the Gulf of Aqaba. We consider it a form of C. textile intergrading with typical C. textile and form neovicarius (see below). - C. textile var. euetrios (Pl. 68, Figs. 2, 3): Last whorl ventricosely conical to ovate, rather narrow (RD 0.53-0.61); spire comparatively high (RSH 0.14-0.20). Ground-colour white, suffused with brownish beige, violet and/or blue. Colour pattern (may be faded in the holotype) close to that of C. t. suzannae and C. t. form archiepiscopus. Shells from Réunion with a blue ground have been referred to as C. t. form euetrios var. cyanosus (Lauer, 1987) (Pl. 68, Fig. 3). Form euetrios is reported from various localities in the W. Indian Ocean. Exact data on its distribution are missing on account of confusion with form archiepiscopus. The holotype of var. euetrios has a slightly narrower last whorl than the lectotype of C. archiepiscopus. - C. eumitus (Pl. 67, Fig. 19): See C. cholmondeleyi. - C. cholmondeleyi f. fascifer: See C. cholmondeleyi. - C. t. neovicarius: (Pl. 67, Fig. 7): Shell rather solid with a usually ventricosely conical last whorl and a rather typical C. textile pattern. Known from the Red Sea and the Gulf of Aden. A form of C. textile intergrading with typical C. textile and form dahlakensis in different localities of its geographical range. - C. textile var. ponderosa: "Form très courte, obèse et lourde" (Dautzenberg). A synonym of form auriger or form verriculum. - C. pyramidalis (Pl. 67, Figs. 12-15): Last whorl rather narrow (RD 0.60 or less) and conoid-cylindrical to ventricosely conical, with a rather widely meshed pattern; spire comparatively high (RSH 0.13-0.19). Known from Kenya and Madagascar. A form of C. textile intergrading with more typically patterned sympatric variants that may also have cylindrical or ovate last whorls. The lectotype (Pl. 67, Fig. 12) may have an aberrantly high spire. - C. scriptus (Pl. 67, Figs. 9-11): Lighter than other forms of C. textile (see Shell Morphometry). Last whorl ventricosely conical or conoid-cylindrical, sometimes approaching ovate or cylindrical (RD 0.55-0.63; PMD 0.74-0.77). Larval shell of about 2 white whorls; spire comparatively high (RSH 0.13-0.18). Spiral colour bands reduced to rows 'of spots and flecks. Known from St. Brandon, Indian Ocean; we consider it a local race of C. textile. - C. sirventi: A form of C. textile with a ventricosely conical last whorl and a reduced pattern. - C. suzannae (Pl. 67, Figs. 16, 17): Smaller than other forms of C. textile (see Shell Morphometry). Last whorl rather narrow, conoid-cylindrical to ventricosely conical (RD 0.52-0.60; PMD 0.75-0.80); spire rather high (RSH 0.13-0.19). Ground colour white to yellowish orange, often suffused with violet. Pattern consisting of reddish brown reticulated lines and 2-3 overlying weak and interrupted spiral colour bands; within bands, lines often not reticulated but axially undulating and sparsely fusing into broad axial lines. Spiral banding may be emphasized by pale violet background shades underlying only the strongly reticulated spiral zones. Known from Malindi and Robinson Id., Kenya. We consider C. suzannae a local race of C. textile most similar in colour pattern to C. t. form euetrios (see below), also resembling forms archiepiscopus and cholmondeleyi. - C. textilinus (Pl. 68, Figs. 9-13): This taxon seems to refer to the local form of C. textile from the Marquesas. Shells of this population have ventricosely conical last whorls (RD 0.57-0.65; PMD 0.76-0.84) with narrowly reticulated spiral zones alternating with bands of yellowish brown blotches and intermittent groups of larger white tents. The aperture is pinkish violet, distinguishing them from typical C. textile that is otherwise not unequivocally separable. Its radular tooth morphology matches that of typical C. textile. Because shells matching those from the Marquesas Is. are known from Tahiti, Tuamotu Archipelago, W. Samoa, Vanuatu, and some W. Indian Ocean localities, C. textilinus must either be regarded as a valid species (Richard, 1990) or a form of C. textile. We consider the conchological differences not to justify separation at the species level. - C. verriculum (Pl. 67, Figs. 1-4): Shell rather solid, with a ventricosely conical or ovate to broadly ovate last whorl (RD 0.59-0.72 vs. 0.50-0.67 in other forms of C. textile; PMD 0.73- 0.78). Larval shell of about 2 pinkish red whorls, maximum diameter 0.7 mm. Colour pattern of rather typical C. textile arrangement. Known from Mauritius, Mozambique, Madagascar and Sri Lanka. The original figures of C. verriculum (see Pl. 67, Fig. 1) are slightly stouter than the lectotype of form auriger. We consider C. verriculum a form of C. textile intergrading with forms auriger and ponderosa. We consider the following nominal species-group taxa mere synonyms rather than forms of C. textile: - C. auratus - C. dilectus (juvenile specimen) - C. gloriamaris (Röding) - C. gloriamaris Perry - D. t. osullivani - C. panniculus (Pl. 68, Fig. 8) - C. reteaureum - C. tigrinus (often erroneously synonymized with C. canonicus).
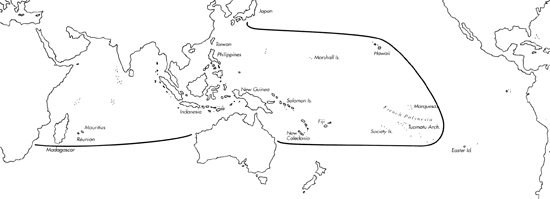
C. textile range map
This section contains verbatim reproductions of the accounts of 316 species of Conus from the Indo-Pacific region, from Manual of the Living Conidae, by Röckel, Korn and Kohn (1995). They are reproduced with the kind permission of the present publisher, Conchbooks.
All plates and figures referred to in the text are also in Röckel, Korn & Kohn, 1995. Manual of the Living Conidae Vol. 1: Indo-Pacific Region.
The range maps have been modified so that each species account has it own map, rather than one map that showed the ranges of several species in the original work. This was necessary because each species account is on a separate page on the website and not confined to the order of accounts in the book.