Range: Indian Ocean except for India; Hawaii.
Description: Medium-sized to large, moderately solid to moderately heavy. Last whorl conoid-cylindrical or ventricosely conical to conical or ovate; outline variably convex at adapical fourth to third, occasionally two-thirds, nearly straight below. Aperture wider at base than near shoulder to uniformly broad. Shoulder angulate to rounded. Spire of low to moderate height; outline concave to convex in late whorls, usually slightly convex to domed in early whorls; apex often straight-sided in shells from S. Madagascar and occasionally in shells from N. Mozambique. In Hawaii, larval shell of about 1.75 whorls, maximum diameter 0.7-0.75 mm; in the Indian Ocean, maximum diameter about 0.8 mm. Teleoconch sutural ramps flat to slightly convex, usually slightly concave in latest whorls; middle ramps with 2 increasing to 3-5 weak spiral grooves, latest ramps usually with numerous spiral striae. Last whorl with weak to distinct ribs basally and with spiral striae from base to shoulder.
| Shell Morphometry | ||
|---|---|---|
| L | - | |
| RW | - | |
| RD | - | |
| PMD | - | |
| RSH | - | |
Ground colour white, often variably suffused with greyish blue to greyish violet, sometimes with yellowish to pinkish cream, orangish to light brown, or with pink to red or violet. Last whorl usually overlaid with yellow or orange to brown, red-brown or blackish brown, leaving many tiny to moderately large tentlike ground-colour markings. Tents separate or fused, edged with a darker line at the frontal sides and usually concentrated in 3 spiral bands, below shoulder, near centre and at base, usually also grouped in axial clusters to form 2-5 axial bands. Dark overlying spots, flecks, and blotches largely solid to sometimes perforated, often forming 2-3 interrupted to continuous spiral bands, and usually with closely spaced, dotted to solid darker spiral lines variably articulated with small ground-colour markings. Shells with a regularly arranged network of fine to coarse dark lines intergrade with shells 1) with a sparsely reticulated, fine or coarse, axially lineate pattern, 2) having last whorl solid dark except for scattered small groups of ground colour tents, and 3) lacking any overlying pattern elements. Basal part of columella white, occasionally violet. Larval whorls and first 1-5 postnuclear sutural ramps pink to purple, sometimes also orange or white; all these colours sometimes occurring among the hatchlings of the same egg capsule but usually one colour preponderant within a local population (Perron, 1981a). Following sutural ramps matching last whorl in colour pattern. Aperture white, to light blue, pink or light violet in some populations.
Periostracum greyish to brownish yellow, thin, translucent, and smooth; rather brown, thicker, less smooth, and opaque in form lohri.
Dorsum of foot white, mottled with brown. Sole of foot mottled light and darker brown. Tentacles white, mottled with brown. Siphon white, tipped with orange-red to red and mottled with brown proximally, with a black ring about 1/3 of the length from the tip (Indian Ocean: Kohn, unpubl. observ.; Fainzilber et al., 1992; Hawaii: Kohn, 1959a). In form lohri, animal white; foot mottled with interlaced brown to blackish brown markings; rostrum grading to beige distally, with blackish brown transverse lines; tentacles tipped with red; siphon matching those of other forms in colour pattern (Ramalho, unpubl. observ.). The colour photograph identified as C. lohri in Liltved and Millard (1989) seems to show an animal of the C. planorbis/C. ferrugineus complex.
Radular teeth with a laterally inflated adapical barb opposite a larger second barb; serration external, extending 1/3 of the length down the shaft; a distinct cusp located about 1/3 of the length from the base; waist and basal spur absent (James, 1980; Bandel, 1984; Nybakken, 1990). Form lohri with very similar radular teeth; serration of a double row of denticles terminating in a cusp (Kilburn, 1972). Subadult specimens (L 9.6 mm) have the same tooth morphology as adult animals. This is correlated with a similar diet (molluscs) in both stages (Nybakken, 1990).
Habitat and Habits: Indian Ocean: From the infralittoral fringe to about 50 m; most frequently on subtidal coral reef flats in 0.5-5 m of water, in coral rubble, sand and muddy sand, often under rocks and amongst or under living corals (Kohn, 1961b, 1968b; Kohn & Nybakken, 1975; Grosch, pers. comm., 1989 & 1990; Fainzilber et al., 1992; Lorenz, pers. comm., 1993). Form lohri reported from 30 m and deeper in Natal (Liltved & Millard, 1989); in S. Mozambique, this form in 2-20 m of water, on rocky ledges with algal turf (Kilburn, 1972; Ramalho, pers. comm., 1989). Hawaii: Rarely on intertidal benches, usually on subtidal reef flats to about 20 m, in sand, on reef rock and coral rubble; animals buried in sand under rocks during day, very often crawling on bare reef limestone at night (Kohn, 1959a, b). C. pennaceus feeds on prosobranch and opisthobranch gastropods but has not been observed to prey on congeners (Kohn, 1959b; Kohn & Nybakken, 1975; Ramalho, pers. comm., 1989). In N. Mozambique, animals known to consume dead fishes, worms, octopods and shrimps (Grosch, pers. comm., 1989). Egg capsules deposited in clusters attached to hard substrate by a few capsules; subsequent capsules affixed to the previously laid forming "arches and bridges". Clusters containing about 60 capsules of 14-17 x 10-11 mm in the Maldives and about 35 capsules of 8.5-13 x 7-10 mm in Hawaii. Observed number of eggs per capsule is 480-660 in Maldives and 25-250 in Hawaii. In the Indian Ocean, egg diameter of 375-407 µm predicts a minimum pelagic period of about 7 days; in Hawaii, egg diameter is 470-520 µm and hatchling is a veliconcha metamorphosing within one day (Ostergaard, 1950; Kohn, 1961a, b; Perron, 1981a, b, c, 1982; Perron & Kohn, 1985).
Discussion: C. pennaceus is similar to C. magnificus, C. episcopatus, C. omaria, C. madagascariensis, and C. echo. C. magnificus generally has a higher spire (RSH 0.13-0.19). Those forms of C. pennaceus with a similarly fine tentmark pattern can be distinguished by their more angulate shoulders and their usually broader last whorls (RD 0.57-0.68 vs. 0.49-0.60), with a conical to ventricosely conical rather than conoid-cylindrical shape. Those forms of C. pennaceus that are similar in shape differ in their coarser pattern with a smaller portion of very small ground-colour tents. Slight differences in the number of larval whorls and the colouration of the animals between C. magnificus from the Marshall Is. And C. pennaceus from Hawaii support the separation at the species level. C. episcopatus is a somewhat larger species that differs in having a generally narrower and more cylindrical last whorl (RD 0.42-0.58; PMD 0.71-0.84) as well as a coarser pattern with a prominent axial arrangement and without darker brown demarcation lines at the frontal sides of its usually larger tents. The distinctive dark and light axial zones of the last whorl pattern in subadult C. episcopatus excludes confusion with similarly sized C. pennaceus. C. omaria differs in having a generally narrower and rather cylindrical last whorl (RD 0.45-0.60 vs. 0.50-0.71) and a yellowish cream to yellow or pinkish orange aperture; the brown spots, flecks, and blotches of its last whorl pattern are interrupted rather than largely solid as in most specimens of C. pennaceus. C. madagascariensis can be distinguished by its consistently straight-sided apex and its rather uniformly reticulated pattern. The difference between C. madagascariensis and C. pennaceus in egg diameters (654 vs. 375-407 or 470-520 µm) strongly suggests separation at the species level, the more so because the greatest difference occurs between nearby populations (C. madagascariensis from S. India and C. pennaceus from Maldives). For comparison with C. echo, see the Discussion of that species. C. pennaceus is so variable in shape and colour pattern that the many nominal species-group taxa do not cover the entire conchological divergence within and between local populations of this species. While most of the Indian Ocean populations are rather uniform in their respective shell characters, a wide range of variation exists in N. Mozambique and Hawaii. In Hawaii, variants differing in colour pattern, shape and solidity of the shell were found among the offspring of the same female (Perron, 1979). We provisionally regard the different local populations from the Indian Ocean as forms. The marginal population from Hawaii perhaps deserves subspecies status, based on the correlative differences in egg diameter and development (benthic in Hawaii, planktonic in the Indian Ocean). Geographical survey of the different forms of C. pennaceus: N. Mozambique (Pl. 63, Figs. 4-12, 14-16): Shape and colouration covering nearly the entire range of variation observed in C. pennaceus. Shells with a fine reticulate pattern including rather small and strongly perforated colour zones intergrade with shells with a fine axially lineate pattern as well as with shells with axially fused ground-colour blotches. The lectotypes of C. pennaceus (Pl. 63, Fig. 1), C. praelatus (Pl. 63, Fig. 3), and the original figure of C. elisae (Pl. 63, Fig. 13) are matched by specimens from N. Mozambique (see Pl. 63, Figs. 4, 5, 11, 12). Shells corresponding with the lectotype of C. elisae in the fine dark axially lineate pattern occur in colonies that also include shells with a partially reticulate and partially lineate pattern. These colonies live sympatrically and sometimes almost syntopically with colonies of shells that have a rather typical pattern on a pale blue to pale violet ground (Pl. 63, Fig. 14; erroneously referred to as "C. praelatus"; Grosch, pers. Comm., 1989 & 1990). The lectotype of C. colubrinus (Pl. 63, Fig. 2) may also be from this area. S. Mozambique, Natal: In the local population of C. pennaceus from Bazaruto Is. (form bazarutensis; Pl. 63, Fig. 17), the last whorl is usually ventricosely conical and has a rather fine and sometimes reduced network of brown to blackish brown lines, spots and blotches on a greyish blue to greyish violet ground; the apex is slightly convex to domed in outline and usually white. Shells very similar in shape and pattern live farther south in the Massinga area. They have a pinkish cream ground colour, a narrow paler spiral band below centre, and a violet apex; their brown reticulate pattern may be more reduced than in form bazarutensis (Pl. 63, Figs. 18,19). Specimens from Natal and S. Mozambique as far north as Inhambane were described as C. lohri ( Pl. 63, Figs. 20-22) but cannot be separated from C. pennaceus from Bazaruto Is. And Massinga by shell morphology and sculpture, nor do they differ from C. pennaceus in body colouration or structure of radular teeth. The report of a multispiral larval shell in C. lohri (Kilburn, 1972) appears doubtful. The ground colour is orangish brown to brown or greyish blue to greyish violet, and sometimes all these colours merge on the same shell. The last whorl often has a broad darker spiral band on each side of centre and occasionally remnants of a brown to blackish brown pattern similar to that of form bazarutensis. The aperture is pale blue to pale violet, edged with a violet collabral band. The low spire has a sigmoid outline and a reddish violet to purple apex. The differences between C. lohri and C. pennaceus in colour, structure and translucency of the periostracum do not allow separation at the species level. Tanzania, Kenya: Last whorl ventricosely conical to conical. Ground colour white or suffused with reddish pink. Shells with strongly perforated brown zones and a fine brown network between intergrade with shells with large solid brown zones and irregularly arranged ground-colour tents between (Pl. 64, Figs. 1-3). Red Sea: Last whorl ventricosely conical to conical; spire usually of moderate height. Pattern consisting of brown to dark brown lines edging small to moderately large white tents; interspersed yellowish brown to dark brown spots and flecks solid but sparse (Pl. 64, Fig. 13). S. Madagascar: In the local form described as C. p. behelokensis, last whorl ventricosely conical to sometimes conoid-cylindrical; early postnuclear whorls with straight to slightly convex sides. Ground colour greyish blue to greyish violet. Shells overlaid with olive, yellowish or orangish brown except for scattered groups of rather small ground-colour tents intergrade with shells with separate brown blotches and spirally as well as axially grouped tents between. Darker brown spiral lines prominent, dotted to solid, and variably articulated with small ground-colour markings (Pl. 64, Figs. 14, 15). Réunion: In the local form described as C. rubropennatus, last whorl ventricosely conical to conoid-cylindrical or conical, ground colour red to reddish violet, and last whorl pattern generally composed of very small to small and regularly arranged ground-colour tents (Pl. 63, Fig. 17). Mauritius, Rodriguez Id.: A local form described by Hwass as C. episcopus is characterized by a comparatively light shell with a ventricosely conical to ovate last whorl. The colour is brown to blackish brown, and the very small to moderately large white tents may be regularly arranged on the last whorl or have larger tents concentrated below shoulder, at centre and at base (Pl. 63, Figs. 23-25). Form episcopus occurs in less than 10 m. Mauritius, St. Brandon: A local form described as C. rubiginosus by Hwass and as C. episcopus mauritiensis by Lauer has a comparatively light shell with a usually conoid-cylindrical to ventricosely conical last whorl. Its colour varies from orange to red-brown. Shells with small to medium-sized ground colour tents regularly arranged on the last whorl intergrade with shells with a coarse axially lineate pattern of larger tents grouped in axial bands or fused into narrow axially elongate blotches (Pl. 63, Figs. 26-29). The lectotypes of C. episcopus and C. rubiginosus are rather similar but specimens of both forms may also differ in shape and pattern. In Mauritius, form rubiginosus occurs in deeper water. Seychelles, Saya de Malha: Shells comparatively solid, with a rather broad, conical to ventricosely conical last whorl. Colour brown to dark brown; pattern consisting of small to moderately large white tents partially grouped in axial and spiral bands (Pl. 64, Figs. 7-9). C. omaria var. marmoricolor (Pl. 64, Fig. 4) is based on a variant of C. pennaceus that may be from this area. Oman, Hormuz: The Oman form described as C. quasimagnificus is comparatively large and heavy and has a rather broad and usually conical last whorl. Its ground colour varies from white to greyish blue. Shells with 2 nearly continuous, light to reddish or blackish brown spiral bands and 3 spiral zones of very small to small ground-colour tents on the last whorl intergrade with shells with separate brown flecks and blotches and spirally as well as axially grouped larger tents between (Pl. 64, Fig. 10, 11). Specimens from Hormuz (Pl. 64, Fig. 12) are somewhat smaller and more ventricose; their pattern has uniformly small and rather regularly arranged ground-colour tents. Maldives, Laccadives: The local form was described as C. p. ganensis. It is characterized by a low spire, an angulate shoulder and a conical last whorl; its colour is yellowish brown to mostly reddish brown (Pl. 64, Fig. 18). Sri Lanka, Andaman Sea, Indonesia: Shells from these marginal Indian Ocean populations have ventricosely conical to conical last whorls, low spires, and a dark reddish brown colour; their medium-sized to moderately large ground-colour tents are separate rather than prominently grouped (Pl. 64, Figs. 5-6, 19). They also may represent form marmoricolor. W. Australia: We have seen 2 specimens from Ashmore reef, N. W. Australia; their last whorl is conical, their spire of moderate height and their pattern includes large overlapping white tents (Pl. 64, Fig. 16). Hawaii (Pl. 64, Figs. 25-28): Shells variable in shape and pattern between and within local populations, usually less variable in colour (various shades of brown). Largest and broadest specimens of C. pennaceus collected in Hawaii (Kohn & Weaver, 1962). Last whorl ventricosely conical to conical or ovate. Ground colour white to greyish blue. Shells with very small to moderately large ground-colour tents, regularly arranged or spirally and axially grouped, intergrade with shells with fine, closely spaced axial brown lines and usually 2 continuous brown spiral bands on the last whorl. Shells with an axially lineate pattern (Pl. 64, Figs. 25) represent an individual variant (Perron, 1979); they are often but probably erroneously referred to as "C. sindon" (see C. omaria, and Pl. 62, Figs. 19-21). Another taxon sometimes applied to the Hawaiian shells is C. stellatus Kiener. However, the original figure (Pl. 64, Fig. 24) shows a subadult shell that cannot be unequivocally assigned to a particular geographic region. The type locality of C. racemosus Sowerby III (Pl. 64, Fig. 23) is given as "Sandwich Islands?", and its holotype resembles Hawaiian specimens in shape. However, the colour pattern must be regarded as very unusual for this area and rather corresponds with that of form behelokensis from Madagascar.
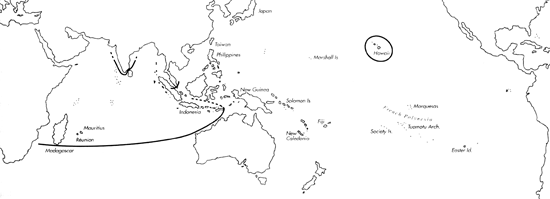
C. pennaceus range map
This section contains verbatim reproductions of the accounts of 316 species of Conus from the Indo-Pacific region, from Manual of the Living Conidae, by Röckel, Korn and Kohn (1995). They are reproduced with the kind permission of the present publisher, Conchbooks.
All plates and figures referred to in the text are also in Röckel, Korn & Kohn, 1995. Manual of the Living Conidae Vol. 1: Indo-Pacific Region.
The range maps have been modified so that each species account has it own map, rather than one map that showed the ranges of several species in the original work. This was necessary because each species account is on a separate page on the website and not confined to the order of accounts in the book.