Range: C. n. nielsenae: Queensland to New South Wales and Kermadec Is.; C. n. reductaspiralis: W. Australia, N. W. Cape to Cape Leveque (Coomans & Filmer, 1985; Turnbull, pers. comm. 1987); C. n. typhon: E. Transkei (South Africa) to Mozambique.
Description: C. n. nielsenae (Pl. 60, Figs. 1, 2): Medium sized, moderately solid. Last whorl conical, occasionally slightly pyriform; outline variably convex at adapical fourth, straight to slightly concave below. Shoulder angulate to slightly carinate. Spire usually low; outline deeply concave, with a conoid apex projecting from an otherwise rather flat spire. First 3-5 postnuclear whorls weakly tuberculate, late whorls often carinate. Teleoconch sutural ramps nearly flat, with 1-2 increasing to 3-5 weak spiral grooves in early whorls and an obsolete spiral striation in late whorls. Basal fourth of last whorl with a few spiral ribs anteriorly and very few spiral ribbons above. C. n. reductaspiralis (Pl. 60, Figs. 3-8): Moderately small to medium sized or rarely to moderately large; moderately solid to solid; in large specimens, C. n. reductaspiralis twice as heavy as C. n. nielsenae. Last whorl usually conical, generally broader than in C. n. nielsenae, particularly in form thevenardensis (Pl. 60, Fig. 8); outline almost evenly straight or grading to convex at adapical fourth to third. Shoulder sharply angulate to carinate, less so in large specimens. Spire of low to moderate height in typical form, usually low in form thevenardensis; outline slightly to deeply concave. Larval shell of about 2 whorls, maximum diameter 1-1.1 mm. Teleoconch spire and last whorl match those of C. n. nielsenae in sculpture. C. n. typhon (Pl. 60, Figs. 9-11): Moderately small to large, moderately solid to solid; deep-subtidal shells may have twice the maximum size of upper-subtidal shells. Last whorl conical to slightly pyriform; outline variably convex at adapical third, straight to slightly concave below. Shoulder carinate, angulate in larger specimens. Spire low, outline usually deeply concave. Larval shell of 1.75-2.25 whorls, maximum diameter 1.1-1.2 mm. Teleoconch spire and last whorl match those of C. n. nielsenae in sculpture except for slightly more pronounced spiral striae on late sutural ramps.
| Shell Morphometry | ||
|---|---|---|
| L | - | |
| (-C. n. nielsenae 40- 55 mm;-C. n. reductaspiralis 27- 51 mm, rarely to 61 mm;-C. n. typhon 30- 91 mm) | ||
| RW | - | |
| (-C. n. nielsenae 0.14- 0.26 g/mm;-C. n. reductaspiralis 0.06- 0.50 g/mm;-C. n. typhon 0.14- 0.41 g/mm L 60-47 mm) | ||
| RD | - | |
| (-C. n. nielsenae 0.56- 0.63;-C. n. reductaspiralis 0.56- 0.70;-f. thevenardensis 0.63- 0.71;-C. n. typhon 0.58- 0.65) | ||
| PMD | - | |
| (C. n. nielsenae 0.90- 0.95;C. n. reductaspiralis 0.85- 0.91;C. n. typhon0.85- 0.94) | ||
| RSH | - | |
| (C. n. nielsenae 0.05- 0.14;C. n. reductaspiralis 0.05- 0.20;-f. thevenardensis 0.04- 0.14;C. n. typhon 0.05- 0.12) | ||
C. n. nielsenae: Ground colour white, merged with light violet. Last whorl variably overlaid with orange, with variably prominent, closely spaced, orange-brown spiral lines. Base without spiral lines and usually lighter than adjacent area. Postnuclear sutural ramps with orange-brown radial streaks extending beyond outer margins. Aperture white, sometimes shaded with violet. Periostracum thin, translucent, with spiral rows of hairlike tufts (Marsh, 1962). C. n. reductaspiralis: Ground colour white. Last whorl immaculate white or more often variably overlaid with yellow to brown and sometimes additionally with violet. Latter shells with closely spaced brown spiral lines extending from shoulder to basal fifth; base light to dark brown, usually darker than adjacent area. Spiral lines often variably reduced, sometimes absent. Larval whorls white to cream, sometimes pale violet. Postnuclear sutural ramps immaculate white to heavily maculated with light to dark brown radial markings; early ramps sometimes immaculate pinkish orange to violet. Aperture white, sometimes violet. Periostracum olive-grey to brown, thin to thick, opaque, with rather closely spaced, slightly tufted spiral ridges on last whorl (Pt. Hedland; Cape Keraudren; Thevenard Id.). C. n. typhon: Ground colour white, sometimes largely suffused with yellow or violet on last whorl. Last whorl with 2 variably broad, light brown or violet spiral bands leaving ground colour zones at base, at centre and sometimes also below shoulder. Closely spaced brown spiral lines extending from base to shoulder, usually reduced at anterior end, sometimes completely absent. Base generally lighter than adjacent area. Immaculate white shells intergrade with heavily patterned shells. Larval whorls white or pale grey to brownish violet. Postnuclear sutural ramps variably suffused with light brown and violet, particularly in early whorls, immaculate or with sparse to numerous brown radial markings. Aperture white, sometimes with a pale violet band on each side of centre. In shells from the upper subtidal, periostracum brown, thin, translucent, and smooth; deep-subtidal shells with a thicker and opaque periostracum bearing widely spaced tufted spiral ridges on the last whorl (Fernandes & Monteiro, 1992).
In C. n. reductaspiralis, animal pale cream flecked with brownish grey and brown (Whitehead, 1967; Singleton, 1979).
Radular teeth with an adapical barb opposite a narrow blade; weak serration from the posterior end of the barb to the posterior end of the blade; base with a spur. In form thevenardensis, serration slightly longer and consisting of 2-3 rows of stronger denticles. Radular teeth of C. n. typhon correspond with those of typical C. n. reductaspiralis in shape and armature (da Motta, 1987b; Kilburn, 1975; Rolán, pers. comm., 1992).
Habitat and Habits: C. n. nielsenae: In 30-100 m, in clean sand (Coomans & Filmer, 1985). C. n. reductaspiralis: Intertidal to about 30 m; on intertidal flats of mud or muddy sand, on clean sand subtidally (Whitehead, 1967; Singleton., 1979; Turnbull, pers. comm., 1987). The population from Thevenard Id. is reported from the infralittoral fringe to 3 m, on muddy sand bottoms of coral reef (da Motta, 1987b; Gabelish, pers. comm., 1989). C. n. reductaspiralis may be active at bright daylight (Singleton, 1979), and one thousand specimens have been observed spawning in tidal creeks north of Pt. Hedland (Turnbull, pers. comm., 1987). C. n. typhon: In 12-120 m. Populations with moderately small to medium-sized shells in 12-70 m on mud bottom; a population with moderately large to large shells was recently discovered in about 120 m off Quissico, S. Mozambique (Fernandes & Monteiro, 1992).
Discussion: C. n. nielsenae and C. n. typhon are similar to C. voluminalis. The latter species can be distinguished by its higher and more massive, projecting adapical part of the spire. Large specimens of C. voluminalis have an outwardly curved shoulder edge; typically patterned shells differ additionally in their long dark axial streaks as well as their more widely spaced and less uniform spiral lines on the last whorl. With regard to the allopatric occurrence of C. nielsenae, C. n. reductaspiralis, and C. typhon, the conchological differences between them do not justify separation at the species level, but suggest the status of geographical subspecies (see Fernandes & Monteiro, 1992). C. thevenardensis was described as a distinct species, although it cannot be distinguished from completely white shells of C. n. reductaspiralis. The differences in the armature of radular teeth are very slight, and it is not known whether they are constant or not. In agreement with Coomans & Filmer (1985) we assign C. thevenardensis to C. n. reductaspiralis as the local form from Thevenard Id.
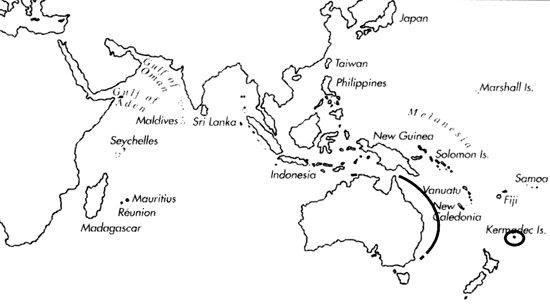
C. nielsenae range map
This section contains verbatim reproductions of the accounts of 316 species of Conus from the Indo-Pacific region, from Manual of the Living Conidae, by Röckel, Korn and Kohn (1995). They are reproduced with the kind permission of the present publisher, Conchbooks.
All plates and figures referred to in the text are also in Röckel, Korn & Kohn, 1995. Manual of the Living Conidae Vol. 1: Indo-Pacific Region.
The range maps have been modified so that each species account has it own map, rather than one map that showed the ranges of several species in the original work. This was necessary because each species account is on a separate page on the website and not confined to the order of accounts in the book.